
Abstract
Despite similar physical properties, the Northern and Southern Atlantic subtropical gyres have different biogeochemical regimes. The Northern subtropical gyre, which is subject to iron deposition from Saharan dust1, is depleted in the nutrient phosphate, possibly as a result of iron-enhanced nitrogen fixation2. Although phosphate depleted, rates of carbon fixation in the euphotic zone of the North Atlantic subtropical gyre are comparable to those of the South Atlantic subtropical gyre3, which is not phosphate limited. Here we use the activity of the phosphorus-specific enzyme alkaline phosphatase to show potentially enhanced utilization of dissolved organic phosphorus occurring over much of the North Atlantic subtropical gyre. We find that during the boreal spring up to 30% of primary production in the North Atlantic gyre is supported by dissolved organic phosphorus. Our diagnostics and composite map of the surface distribution of dissolved organic phosphorus in the subtropical Atlantic Ocean reveal shorter residence times in the North Atlantic gyre than the South Atlantic gyre. We interpret the asymmetry of dissolved organic phosphorus cycling in the two gyres as a consequence of enhanced nitrogen fixation in the North Atlantic Ocean4, which forces the system towards phosphorus limitation. We suggest that dissolved organic phosphorus utilization may contribute to primary production in other phosphorus-limited ocean settings as well.
Similar content being viewed by others

Main
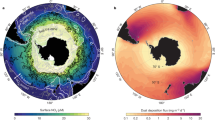
The physical characteristics of the extensive subtropical gyres restrict the supply of inorganic nutrients to surface waters, producing an oligotrophic environment5. Nitrogen (N) and phosphorus (P) are essential micronutrients for oceanic primary production and occur in the ratio of ∼16:1 (ref. 6). However, comparisons of global ocean nitrate and phosphate show that nitrate is exhausted before phosphate7. Thus, in oligotrophic systems where inorganic nutrients are depleted and nitrate is exhausted, a residual phosphate pool should remain. There is, however, a clear contrast between the North Atlantic subtropical gyre (NASG) and the South Atlantic subtropical gyre (SASG: Fig. 1a), where our recent field measurements during seven cruises show contrasting values of surface (25 m depth) phosphate, with mean values of 9 and 210 nM, respectively. This contrast is probably due to the greater role of nitrogen fixation in the NASG (ref. 4) (Fig. 2a), providing an alternative source of N and driving complete draw-down of the residual phosphate pool.

a, 25 m phosphate concentrations (nM) and AMT16 (dashed line) and AMT17 (solid line) ship transects. b, 25 m DOP concentrations (nM). Stations are marked as black dots.

a, Meridional variations in δ15PONsusp above the nitricline (values averaged) for cruises AMT12, AMT14, AMT16 and AMT17 (ref. 4), with depleted values of δ15PONsusp indicating N2 fixation. b, Phosphate concentrations from AMT16 (June; dashed line) and AMT17 (November; solid line). Precision and reproducibility are shown as error bars (±3%). c, APA activities normalized to total carbon. Error bars of duplicate measurements are shown as ±percentage range.d, DOP concentrations. Reproducibility is shown as error bars (±7%).
A possible alternative source of P to phytoplankton in the NASG is dissolved organic phosphorus (DOP). Our observational surveys, incorporating data from seven cruises between April 2000 and November 2005, show that in the surface (25 m depth) of the oligotrophic Atlantic, DOP represents a large fraction of total dissolved phosphorus of 66±23% (s.d.) (see Supplementary Information, Table S1, for individual cruise transects and dates). There are significantly lower absolute concentrations of DOP in the NASG (80±140 nM (s.d.)) than in the SASG (150±70 nM (s.d.)), where the distribution is more homogenous (Fig. 1b). This gyre-scale contrast in the mean DOP is a robust signal and statistically significant, with a concentration±standard error (s.e.m.) of 80±10 nM in the NASG and 150±10 nM in the SASG (s.e.m.=s.d./√(n−1), where s.d. is the standard deviation and n is the number of independent data points; P<0.01; T-test). Given the apparent nutrient asymmetry between the NASG and SASG, we now investigate the role of DOP within the system by examining its turnover and bioavailability in the two gyres, and explore the implications for their biogeochemistry.
The turnover of the DOP pool can be assessed through the measurement of the activity of the P-specific enzyme alkaline phosphatase (APA). This enzyme, which releases P bound in DOP, has been found associated with bacteria and phytoplankton and free in the water8, and its activity can limit DOP decomposition and plankton growth9. Extracellular APA has also been found to be expressed in a large range of phytoplankton taxa in response to P limitation8.
During June and November, the contrasts between three distinct oceanic dynamical regimes were investigated, identified through chlorophyll a and inorganic nutrient concentrations (see the Methods section below): the downwelling NASG (15∘ N–40∘ N), the region of equatorial and wind-induced upwelling (15∘ N–10∘ S) and the downwelling SASG (10∘ S–40∘ S). Michaelis–Menten kinetics were additionally used during spring in each gyre to assess the turnover time and lability of the DOP pool through this season.
Our measured APA activities indicate a significantly greater utilization of the DOP pool in the NASG compared to the SASG over June and November (Fig. 2c). In the NASG, APA activities were ∼3-fold higher in June, with a maximal rate of 4.65 nM P h−1/μg C (mean 2.44 nM P h−1/μg C), compared to a mean of 0.84 nM P h−1/μg C in November. The higher activities correspond to a significantly smaller DOP pool size, which was ∼60% lower in June compared with November (Fig. 2d). These seasonal APA activities and DOP concentrations are significantly different, as indicated by the contrast in their means±standard errors (Table 1; P<0.05; T-test). Phytoplankton community production and growth are higher during spring in both hemispheres3. We interpret the higher APA activities and lower DOP concentrations in the NASG as reflecting phosphate limitation (Fig. 2b), which leads to a greater utilization of the DOP pool to maintain production.
In contrast to the seasonality of the NASG, APA activities of the SASG were lowest in spring (November), with activities of 0.20 nM P h−1/μg C (Fig. 2c). Within these waters, phosphate concentrations were higher in spring (Fig. 2b), likely through winter mixing of the water column. Phosphate is a more bioavailable nutrient source than DOP; hence we observe a reduction in enzyme activities and an accumulation of DOP in the water column (Fig. 2d).
Plots of APA versus phosphate (Fig. 3a) and DOP (Fig. 3b) for the NASG and SASG highlight the differences for spring in the two systems, as there are lower phosphate concentrations in the NASG and subsequently higher APA and lower DOP compared to the SASG. At a confidence level of 98% (P<0.02; T-test) log10 plots of APA versus phosphate and APA versus DOP for all data show significant negative correlations (coefficient of determination r2=0.54 and 0.45, respectively; data not shown). APA activities do not directly represent DOP uptake, as enzyme activities are measured using an artificial substrate present at a higher concentration than the naturally occurring DOP pool. This is probably responsible for the low r2 values reported. It is clear however that the NASG and SASG are biogeochemically different throughout spring owing to the severely depleted levels of phosphate in the NASG.

a, APA activities versus phosphate concentrations. b, APA activities versus DOP concentrations. NASG spring is represented as grey circles, NASG autumn as white diamonds, SASG spring as black triangles and SASG autumn as white squares. Error bars of duplicate APA measurements are shown as±percentage range. Error bars of phosphate and DOP measurements are shown as reproducibility errors (±3% and ±7% respectively).
It is possible to estimate the amount of primary production potentially supported by the DOP pool in spring using the Michaelis–Menten equations. By combining these with the ambient DOP concentrations we can calculate the in situ APA activity. Integrating these activities over the euphotic zone shows the DOP pool to contribute 0.015±0.005 mol P m−2 y−1 to the NASG in spring compared with 0.003±0.001 mol P m−2 y−1 in the SASG. Combined with euphotic-zone rates of carbon fixation for the same season3, and applying the Redfield ratio, we estimate the DOP pool to support 20% (range of 12–30%) of production in the NASG and only 5% (range of 3–7%) in the SASG. Estimates of production in the NASG do vary, however; for example, geochemical estimates of new production in the Sargasso Sea show the P demand to be 0.031±0.009 mol P m−2 y−1 (ref. 10). Using this estimate, the DOP pool would support up to 90% of production.
Given the different utilization rates of DOP in each gyre in spring, we now consider the biological availability of the DOP pool using the Michaelis constant (Km). Bulk dissolved organic matter is often viewed as a series of pools, representing a continuum of biological lability, from refractory material turning over on timescales of centuries to millennia, to very labile material turning over on timescales of minutes to days11. Km provides a measure of the binding strength between the enzyme and DOP; low Km values indicate that labile DOP is scarce and that it is strongly bound by enzymes.
In spring between the NASG (June) and the SASG (November) the variations in Km are not significantly different, with a mean±s.e.m. of 789±387 μM (n=3) and 565±114 μM (n=10) respectively (see Supplementary Information, Table S2 for individual station locations). The high Km values reveal the presence of similar pools of labile DOP in both gyres during spring, presumably due to increased production resulting in the release of fresh, labile DOP compounds, via excretion, exudation, grazing and cell lysis12.
The turnover time of the DOP pool (T) is diagnosed using the Li parametrization, equation (1) (ref. 13), from the ratio of Km and Vmax, the measured activity (μM h−1):

The turnover times of the DOP pools are only 5.5±2.3 months (s.d.) in the NASG, but much longer in the SASG with 10.5±6.8 years (s.d.). In turn, these different turnover rates between the northern and southern gyres imply contrasting DOP distributions over the NASG and SASG. Model studies reveal that high concentrations of DOP produced in the tropics may be transported into the subtropical gyre as part of the wind-driven Ekman and gyre circulation14. A typical advective timescale for a transient tracer to cross the entire NASG within the upper thermocline is 7–10 years (ref. 15). Consequently, short DOP turnover times of less than a year in the NASG imply that there is insufficient time for the DOP to be transported over the entire gyre, resulting in consistently lower concentrations within the interior of the gyre as seen in the observations (Fig. 1b). Conversely, in the SASG, the DOP turnover times of typically 10 years are sufficiently long for DOP to be advected over the entire gyre and become relatively uniform.
These experimental diagnostics reveal opposing biogeochemical regimes in the NASG and the SASG, with the DOP pool acting as an important nutrient source to marine plankton in the NASG as a result of severely depleted phosphate concentrations. The NASG is documented for its atmospheric, synoptic-scale dust inputs and resulting deposition of iron1. Studies of the distribution of the stable isotopes of nitrogen in suspended particulate organic matter likewise reveal a depleted signal over the central and eastern side of the NASG, between 10∘ N and 30∘ N, consistent with regions of N2 fixation4 (Fig. 2a). Furthermore, the nitrogen-fixing cyanobacteria Trichodesmium spp. occur extensively within the warm waters of the North Atlantic16, are uniquely adapted for scavenging P from organic sources17 and have been shown to contribute substantially to total APA in the water18. Viewed together, these observations suggest that N2 fixation leads to the observed depleted surface phosphate concentrations and subsequent raised levels of APA activity and DOP utilization in the NASG. The opposing biogeochemistry of the NASG and SASG probably reflect marked differences in the atmospheric delivery of dust, implying that this asymmetry might vary with the ongoing, long-term changes in the atmospheric winds19.
The contrast in P cycling has wider significance in the ocean export of organic carbon, because half the global export of organic carbon is estimated to occur over these extensive, oligotrophic subtropical gyres20. This export of organic matter is sustained through a supply of new nutrients to the sunlit, surface ocean, which we show over the NASG to be partly achieved through the enhanced utilization of DOP. The asymmetry of the P cycling in the NASG and SASG probably reflects the enhanced levels of nitrogen fixation in the NASG, forcing4 the system towards P limitation. The enhanced utilization of DOP in the NASG might therefore also be relevant to other extensive, oligotrophic P-limited parts of the global ocean, such as the Mediterranean Sea21 and, possibly, the North Pacific subtropical gyre22. Thus, utilization of DOP potentially sustains levels of carbon fixation over much of the global oligotrophic ocean whenever there is phosphorus limitation.
Methods
Seven cruises were conducted between April 2000 and November 2005. Seawater samples were collected from Niskin bottles mounted on a SeaBird CTD/Rosette system.
Alkaline phosphatase activities
APA activities in the photic zone were determined at a total of 37 stations along two Atlantic meridional transects (AMTs) in boreal spring (AMT16; 19 May–29 June, referred to as June) and autumn in 2005 (AMT17; 15 Oct–28 Nov, referred to as November; Fig. 1a). Reported activities were normalized to biomass. The fluorogenic substrate 4-methyllumbelliferylphosphate was used to assess the alkaline phosphatase activities, using substrate additions of 250 μM for AMT17 and 200 μM for AMT16. Michaelis–Menten kinetics were used for the NASG in June and the SASG in November on cruises AMT16 and AMT17. The key parameters from these experiments were Vmax, the maximum rate of turnover of DOP by the enzyme, and Km, the concentration of DOP at which the enzymes are half saturated. Michaelis–Menten kinetic experiments were conducted with three to six 10 ml seawater samples inoculated with 1–750 μM substrate, in acid-washed polypropylene pots. All experiments were incubated in the dark, at in situ temperature, for 24–48 hours. Details of station experiments and locations are included in the Supplementary Information, Table S2, which accompanies this paper.
Normalization
Activities were normalized to total carbon, assessed through chlorophyll a concentrations and bacterial counts, using conversion factors of 1 g chl a=50 g C (ref. 23) and 12 fg C/bacterial cell (ref. 24). Chlorophyll a concentrations were determined using a fluorometric assay of the acetone extract of particulate material collected on a GF/F filter25. Errors were estimated at ±5%. Bacterial counts were carried out by flow cytometry, using a 15 mW laser set to excite at 488 nm (ref. 26). Errors were estimated at ±2.5%. The patterns we report were found to be significant regardless of the method of normalization used, that is, no normalization, bacterial numbers only, chlorophyll concentrations only or total carbon.
Nutrient concentrations
Phosphate concentrations were analysed using standard colorimetric techniques27,28, with an analytical precision of ±3%. DOP samples from cruises AMT10, CD171 and AMT17 were filtered immediately on collection using ashed GF/F filters and a glass filtration unit, and stored frozen in sterile high-density polythene 60 ml bottles. Samples from cruises D279, AMT14, AMT15 and AMT16 were unfiltered and therefore represent total organic phosphorus, which is appropriate in oligotrophic waters, where the particulate pool is generally considered negligible (<10%) (ref. 29), and has hence been referred to as DOP throughout the text. DOP was calculated as the difference of total dissolved phosphorus and phosphate, measured by standard colorimetric techniques following ultraviolet photo-oxidation30 with a Metrohm 705 ultraviolet digestion system. Reproducibility errors were ±7% on the basis of duplicate measurements from AMT16.
Statistics
Statistical analysis of the data sets was carried out by Excel data analysis (one-tailed T-test) and Sigma plot and Sigma-stat analytical tools (test of normal distribution). See Supplementary Information, Table S3–S7 for individual P values.
References
Duce, R. A. & Tindale, N. W. Atmospheric transport of iron and its deposition in the ocean. Limnol. Oceanogr. 36, 1715–1726 (1999).
Wu, J., Sunda, W., Boyle, E. A. & Karl, D. M. Phosphate depletion in the Western Atlantic Ocean. Science 289, 759–762 (2000).
Poulton, A. J. et al. Phytoplankton carbon fixation, chlorophyll-biomass and diagnostic pigments in the Atlantic Ocean. Deep-Sea Res. II 53, 1593–1610 (2006).
Reynolds, S. E. et al. How widespread and important is N2 fixation in the North Atlantic Ocean? Glob. Biogeochem. Cycles 21, GB4015 (2007).
Williams, R. G. & Follows, M. J. in Ocean Biogeochemistry: The Role of the Ocean Carbon Cycle in Global Change (ed. Fasham, M.) (Springer, Berlin, 2003).
Redfield, A. C. in On The Proportions of Organic Derivatives in Seawater and their Relation to the Composition of Plankton (ed. Daniel, R. J.) (Univ. Press of Liverpool, Liverpool, 1934).
Tyrell, T. The relative influences of nitrogen and phosphorus on oceanic primary production. Nature 400, 525–531 (1999).
Beardall, J., Young, E. & Roberts, S. Approaches for determining phytoplankton nutrient limitation. Aquat. Sci. 63, 44–69 (2001).
Hoppe, H.-G. in Handbook of Methods in Aquatic Microbial Ecology (eds Kemp, P., Sherr, B., Sherr, E. & Cole, J. J.) 423–431 (Lewis Publishers, Boca Raton, FL, 1993).
McGillicuddy, D. J. Jr & Robinson, A. R. Eddy-induced nutrient supply and new production in the Sargasso Sea. Deep-Sea Res. I 44, 1427–1450 (1997).
Carlson, C. A. in Biogeochemistry of Marine Dissolved Organic Matter (eds Hansell, D. A. & Carlson, C. A.) 91–151 (Academic, London, 2002).
Karl, D. M. & Byorkman, K. M. in Biogeochemistry of Marine Dissolved Organic Matter (eds Hansell, D. A. & Carlson, C. A.) 249–367 (Academic, London, 2002).
Li, W. K. W. Consideration of errors in estimating kinetic-parameters based on Michaelis–Menten formalism in microbial ecology. Limnol. Oceanogr. 28, 185–190 (1983).
Roussenov, V., Williams, R. G., Mahaffey, C. & Wolff, G. A. Does the transport of dissolved organic nutrients affect export production in the Atlantic Ocean? Glob. Biogeochem. Cycles 20, GB3002 (2006).
Jenkins, W. J. H-3 and He-3 in the beta-triangle—observations of gyre ventilation and oxygen utilization rates. J. Phys. Oceanogr. 17, 763–783 (1987).
Tyrrell, T. et al. Large-scale latitudinal distribution of Trichodesmium spp. in the Atlantic Ocean. J. Plankton Res. 25, 405–416 (2003).
Dyhrman, S. T. et al. Phosphonate utilization by the globally important marine diazotroph Trichodesmium. Nature 439, 68–71 (2006).
Sohm, J. A. & Capone, D. G. Phosphorus dynamics of the tropical and subtropical North Atlantic: Trichodesmium spp. versus bulk plankton. Mar. Ecol. Prog. Ser. 317, 21–28 (2006).
Hurrell, J. W. in The North Atlantic Oscillation: Climate Significance and Environmental Impact (eds Kushnir, Y., Ottersen, G. & Visbeck, M.) (American Geophysical Union, Washington, DC, 2003).
Emerson, S. et al. Experimental determination of the organic carbon flux from open-ocean surface waters. Nature 389, 951–954 (1997).
Thingstad, T. F. et al. Nature of phosphorus limitation in the ultraoligotrophic eastern Mediterranean. Science 309, 1068–1071 (2005).
Karl, D. M., Bidigare, R. R. & Letelier, R. M. Long-term changes in plankton community structure and productivity in the North Pacific Subtropical Gyre: The domain shift hypothesis. Deep-Sea Res. II 48, 1449–1470 (2001).
Antia, N. J., McAllistel, C. D., Parsons, T. R., Stephens, K. & Strickland, J. D. H. Further measurements of primary production using a large-volume plastic sphere. Limnol. Oceanogr. 8, 166–183 (1963).
Simon, M. & Azam, F. Protein content and protein synthesis rates of planktonic marine bacteria. Mar. Ecol. Prog. Ser. 51, 201–213 (1989).
Welschmeyer, N. A. Fluorometric analysis of chlorophyll-A in the presence of chlorophyll-B and pheopigments. Limnol. Oceanogr. 39, 1985–1992 (1994).
Zubkov, M. V., Sleigh, M. A., Burkill, P. H. & Leakey, R. J. G. Picoplankton community structure on the Atlantic Meridional Transect: A comparison between seasons. Prog. Oceanogr. 45, 369–386 (2000).
Sanders, R. & Jickells, T. Total organic nutrients in Drake Passage. Deep-Sea Res. I 47, 997–1014 (1999).
Woodward, E. M. S. & Rees, A. P. Nutrient distributions in an anticyclonic eddy in the northeast Atlantic Ocean, with reference to nanomolar ammonium concentrations. Deep-Sea Res. II 48, 775–793 (2001).
Abell, J., Emerson, S. & Renaud, P. Distributions of TOP, TON and TOC in the North Pacific subtropical gyre: Implications for nutrient supply in the surface ocean and remineralization in the upper thermocline. J. Mar. Res. 58, 203–222 (2000).
Armstrong, F. A. J., Williams, P. M. & Strickland, J. D. H. Photo-oxidation of organic matter in sea water by ultra-violet radiation analytical and other applications. Nature 211, 481–483 (1966).
Acknowledgements
We thank A. Poulton for the analysis of chlorophyll a, M. Zubkov for the determination of bacterial numbers and K. Chamberlain, T. Lesworth and M. Stinchcombe for help with the analysis of inorganic and organic nutrients. We are grateful to the office, crew and technical support of RRS J. C. Ross, RRS Discovery and RRS C. Darwin. This study was supported by the UK Natural Environment Research Council through the Atlantic Meridional Transect consortium (NER/O/S/2001/00680) and the 36∘ N consortium (NER/O/S/2003/00625).
Author information
Authors and Affiliations
Contributions
R.L.M., S.E.R., G.A.W. and R.G.W wrote the paper. G.A.W. and R.G.W. designed the research programme. R.L.M., S.E.R., S.T.V., E.M.S.W., A.L., X.P., and R.S. carried out the field work. R.S. and E.P.A. oversaw the analysis of the organic nutrients.
Corresponding author
Supplementary information
Supplementary Information
Supplementary tables S1-S7 (PDF 130 kb)
Rights and permissions
About this article
Cite this article
Mather, R., Reynolds, S., Wolff, G. et al. Phosphorus cycling in the North and South Atlantic Ocean subtropical gyres. Nature Geosci 1, 439–443 (2008). https://doi.org/10.1038/ngeo232
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/ngeo232
This article is cited by
-
A dataset of global ocean alkaline phosphatase activity
Scientific Data (2023)
-
Methylphosphonate-driven methane formation and its link to primary production in the oligotrophic North Atlantic
Nature Communications (2023)
-
Seasonal variation in alkaline phosphatase activity in the central part of Tokyo Bay, 2013
Journal of Oceanography (2023)
-
Global and seasonal variation of marine phosphonate metabolism
The ISME Journal (2022)
-
Dissolved organic phosphorus concentrations in the surface ocean controlled by both phosphate and iron stress
Nature Geoscience (2022)
